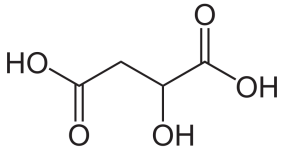
I read a paper the other day claiming that the biological role of mescaline and other related alkaloids is unknown.
Why would an organism devote energy to producing and storing these molecules in such significant amounts?
I believe I have a perspective and an explanation that might be of interest to some here.
Lets start with Tyramine, a well known alkaloid:

It's a hydroxylated phenolic with an amine.
That's easy enough to see.
What about if we methylate that amine?
We call that Hordenine or dimethyltyramine:

It's an N methylated Tyramine, just like the name suggests.
Well, what if we add more to this?
Why not hydroxylate and methylate it a bit more?
Here we have an example of that:

This tyramine molecule has a methoxy group on the 3 position of the molecule!
That's probably why it is called 3-methoxy-tyramine.
In order to do this the organism first adds a hydroxl group to the phenolic ring and then that is O-methylated by a methyltransferase.
In fact, if we do this to the 3, the 4 and the 5 position of the molecule we get 3,4,5-Trimethoxy-Phenyl-ethyl-amine, emphasis on the structural components of PEA being mine. Phenyethylamine or PEA is basically tyramine without any hydroxylation. PEA can be described as being composed of a phenolic group, an ethyl group and an amine.
This is it:

In fact, for cactus alkaloids it is obvious that they proceed from tyramine to other forms, via hydroxylation and methylation specifically.
Something else is worth noticing here.
It has to do with the observations about alkaloid production.
You see, in numerous studies, tests and anecdotes alkaloid+ cacti have specific patterns of alkaloid production and distribution.
Take the dark treatment of cacti, like San Pedro as an example.
The prolonged dark period causes alkaloids to gradually increase and then after the darkness continues they then decrease.
In Peyote studies something similar happens. Drought causes the alkaloids to increase and then after watering the plants the alkaloids then decrease.
In peyote this coincides with a flush of flowers and fruit.
In plants exposed to darkness, after a time, there is an up-regulation of genes associated with the degradation of chlorophyll.
That is to say, plants activity break down their chlorophyl and this allows them to use the materials it is made from elsewhere.
However, alkaloids made do not just stick around, in peyote after watering the amount decreases and in San Pedro in the dark the alkaloids also eventually decrease.
This is potent evidence that the alkaloids are being metabolized and used as sources of energy and molecular feedstock.
There is an indication that methylation and hydroxylation of alkaloids allows plants to store Nitrogen, Oxygen, Carbon and Hydrogen. Essentially by adding and removing hydroxyls and methyl groups a plant can take the same basic subunits that sugar is made from and store them in a form that allows it to be used by the plant, but which prevents that stored energy from being exploited by other organisms. The biology of peyote, where flushes of growth correspond to a decrease in alkaloids make it quite clear that alkaloids are being used as energy and molecular storage methods. This appears to work by hydroxylation and methylation, which appear to be reversible, otherwise alkaloids would never decrease or dissapear once formed. They are not permanent molecules and there is evidence they are also actively transported by plants to tissues with metabolic activity, and that the alkaloids help buffer the CAM acids, which are also formed from the same basic components of carbon, oxygen and hydrogen.
The origin of PEA alkaloids predates the origin of plants and several of the roles that the alkaloids can serve in biology relate to this. Many of them can interfere with cellular processes, this allows them to be energy storage mechanisms that offer protection against ingestion and infection by microbes, fungi, viruses and animals. However no actual selective pressure exists in nature that specifically selects for mescaline over other variations of the molecule, like Pellotine.
Pellotine:

Note that Pellotine has cycled from a PEA to an Isoquinoline via ring closure at the amine, but it is just another collection of methyl and hydroxyl groups on a tyramine skeleton.
This is where things get interesting, as that cacti exist in which mescaline is the dominant alkaloid. In fact as a trace alkaloid mescaline is widespread throughout cacti, but as the primary component of an alkaloid+ cactus it is known only from San Pedro cacti and then only from specific populations, specimens and cases, for many San Pedro cacti contain little to no mescaline or contain modest amounts but have other alkaloids present as the dominant alkaloid.
I am focusing on PEA alkaloids here, but the Tryptamine alkaloids are quite similar in many ways, I may get to them, perhaps not anytime soon.
Alkaloids in cacti allow them to accumulate and store molecular energy in a form resistant to attack. This proceeds by stepwise hydroxylation and methylation and is known to be promoted by conditions known to degrade chlorophyl. The cacti then metabolize this stored energy for growth and reproduction. Many different molecular variations can facilitate this and mescaline is but one of them.
This is a work in progress that I will revise as needed.
Why would an organism devote energy to producing and storing these molecules in such significant amounts?
I believe I have a perspective and an explanation that might be of interest to some here.
Lets start with Tyramine, a well known alkaloid:
It's a hydroxylated phenolic with an amine.
That's easy enough to see.
What about if we methylate that amine?
We call that Hordenine or dimethyltyramine:
It's an N methylated Tyramine, just like the name suggests.
Well, what if we add more to this?
Why not hydroxylate and methylate it a bit more?
Here we have an example of that:
This tyramine molecule has a methoxy group on the 3 position of the molecule!
That's probably why it is called 3-methoxy-tyramine.
In order to do this the organism first adds a hydroxl group to the phenolic ring and then that is O-methylated by a methyltransferase.
In fact, if we do this to the 3, the 4 and the 5 position of the molecule we get 3,4,5-Trimethoxy-Phenyl-ethyl-amine, emphasis on the structural components of PEA being mine. Phenyethylamine or PEA is basically tyramine without any hydroxylation. PEA can be described as being composed of a phenolic group, an ethyl group and an amine.
This is it:
In fact, for cactus alkaloids it is obvious that they proceed from tyramine to other forms, via hydroxylation and methylation specifically.
Something else is worth noticing here.
It has to do with the observations about alkaloid production.
You see, in numerous studies, tests and anecdotes alkaloid+ cacti have specific patterns of alkaloid production and distribution.
Take the dark treatment of cacti, like San Pedro as an example.
The prolonged dark period causes alkaloids to gradually increase and then after the darkness continues they then decrease.
In Peyote studies something similar happens. Drought causes the alkaloids to increase and then after watering the plants the alkaloids then decrease.
In peyote this coincides with a flush of flowers and fruit.
In plants exposed to darkness, after a time, there is an up-regulation of genes associated with the degradation of chlorophyll.
That is to say, plants activity break down their chlorophyl and this allows them to use the materials it is made from elsewhere.
However, alkaloids made do not just stick around, in peyote after watering the amount decreases and in San Pedro in the dark the alkaloids also eventually decrease.
This is potent evidence that the alkaloids are being metabolized and used as sources of energy and molecular feedstock.
There is an indication that methylation and hydroxylation of alkaloids allows plants to store Nitrogen, Oxygen, Carbon and Hydrogen. Essentially by adding and removing hydroxyls and methyl groups a plant can take the same basic subunits that sugar is made from and store them in a form that allows it to be used by the plant, but which prevents that stored energy from being exploited by other organisms. The biology of peyote, where flushes of growth correspond to a decrease in alkaloids make it quite clear that alkaloids are being used as energy and molecular storage methods. This appears to work by hydroxylation and methylation, which appear to be reversible, otherwise alkaloids would never decrease or dissapear once formed. They are not permanent molecules and there is evidence they are also actively transported by plants to tissues with metabolic activity, and that the alkaloids help buffer the CAM acids, which are also formed from the same basic components of carbon, oxygen and hydrogen.
The origin of PEA alkaloids predates the origin of plants and several of the roles that the alkaloids can serve in biology relate to this. Many of them can interfere with cellular processes, this allows them to be energy storage mechanisms that offer protection against ingestion and infection by microbes, fungi, viruses and animals. However no actual selective pressure exists in nature that specifically selects for mescaline over other variations of the molecule, like Pellotine.
Pellotine:
Note that Pellotine has cycled from a PEA to an Isoquinoline via ring closure at the amine, but it is just another collection of methyl and hydroxyl groups on a tyramine skeleton.
This is where things get interesting, as that cacti exist in which mescaline is the dominant alkaloid. In fact as a trace alkaloid mescaline is widespread throughout cacti, but as the primary component of an alkaloid+ cactus it is known only from San Pedro cacti and then only from specific populations, specimens and cases, for many San Pedro cacti contain little to no mescaline or contain modest amounts but have other alkaloids present as the dominant alkaloid.
I am focusing on PEA alkaloids here, but the Tryptamine alkaloids are quite similar in many ways, I may get to them, perhaps not anytime soon.
Alkaloids in cacti allow them to accumulate and store molecular energy in a form resistant to attack. This proceeds by stepwise hydroxylation and methylation and is known to be promoted by conditions known to degrade chlorophyl. The cacti then metabolize this stored energy for growth and reproduction. Many different molecular variations can facilitate this and mescaline is but one of them.
This is a work in progress that I will revise as needed.
Last edited:


.png")