entheogenic-gnosis
Rising Star
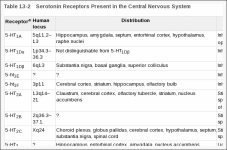
Serotonin Receptors: Genetic insights into serotonin function
Mark J.S. HeathRené Hen
-eg
Mark J.S. HeathRené Hen
The classical pharmacological techniques of using selective agonists, antagonists and inhibitors provided the initial basis for categorizing the receptors, transporters and degrading enzymes involved in serotonergic pathways. But the complexity of the system has overwhelmed the armamentarium of selective drugs, leaving many proteins without specific probes of their function. Recent advances in the field have exploited deliberate or naturally occurring disruptions of the genes that code for proteins important to serotonergic transmission. Four recent contributions [3], [4], [5] and [6] describe the effects of disrupting the genes encoding the 5-HT1B and 5-HT2C receptors in mice, and the monoamine oxidase A (MAOA) genes in mouse and human
5-HT1B knockout — locomotion and aggression
The 5-HT1B receptor was the first protein involved in serotonergic signaling to have its gene disrupted [3]. The 5-HT1B receptor is expressed in the basal ganglia, central gray, hippocampus, amygdala and raphe nuclei; it is targeted primarily to presynaptic terminals, where it can inhibit neurotransmitter release. Highly specific agonists and antagonists are not available for 5-HT1B receptors, but pharmacological experiments suggest that 5-HT1B receptor activation may increase anxiety and locomotion and decrease food intake, sexual activity and aggression. Targeted disruption was achieved by deleting a fragment of the coding sequence, and functional inactivation of the 5-HT1B receptor was confirmed by autoradiography with 125I-cyanopindolol, which binds 5-HT1B receptors, along with appropriate blockers of non-5-HT1B binding sites. Heterozygote animals expressed the same levels of receptor as wild-types, suggesting that a feedback mechanism may regulate the transcriptional activity of the gene.
Two of the behaviors postulated to be modulated by 5-HT1B receptors were analyzed: locomotion and aggression. Untreated wild-type and homozygous mutant (5-HT1B−/−) mice were found to display similar levels of locomotor activity in an open field. However, when treated with the 5-HT1 agonist RU24969, the 5-HT1B−/− mice displayed no change in activity, whereas wild-type mice doubled their activity. This result confirmed previous pharmacological studies indicating that RU24969 operates through 5-HT1B receptors.
The aggressiveness of 5-HT1B−/− male mice was assessed by isolating them for four weeks and then exposing them to a non-isolated male wild-type intruder mouse. The latency and number of attacks displayed by the knockout mice during a three-minute period were used as indices of aggression. The 5-HT1B−/− mice, when compared with wild-type mice, exhibited faster, more intense and more frequent attacks. For example, 46% of the 5-HT1B−/− mice attacked the intruder within 10 seconds of introduction, whereas no heterozygous or wild-type mice attacked during this interval. Tail rattling, an aggressive display, was also more frequent in 5-HT1B−/− mice.
These experiments suggest an involvement of 5-HT1B receptors in the modulation of aggressive behavior. Eltoprazine and fluprazine, drugs classified as serenics because of their anti-aggressive properties, are believed to operate as agonists of 5-HT1 receptors. The lack of subtype-specific antagonists has impeded specific identification of the molecular target of such serenics; the availability of 5-HT1B−/− mice should allow more detailed evaluation of the pathways leading to aggression.
5-HT2C knockout — seizures and overeating
The 5-HT2C gene knockout was generated by insertion of a stop codon that eliminated the carboxyl terminus of the protein [4]. The 5-HT2C gene is X-linked; brains of mutant males were devoid of 5-HT2C immunoreactivity, and mRNA from such males failed to produce functional receptor when injected into Xenopus oocytes.
The patterns of expression of 5-HT2C receptors in the hippocampus and spinal cord suggest that they might modulate memory and nociception. However, hippocampal LTP and nociceptive thresholds were the same in wild-type and 5-HT2C−/− mice. Video monitoring revealed that 5-HT2C−/− mice displayed spontaneous epileptic seizures that were sometimes fatal; survival plots indicated that almost half of the mice had died by 25 weeks of age. Metrazol, a γ-amino butyric acid (GABA) receptor antagonist, was used to quantify seizure susceptibility. Relative to wild-type mice, the 5-HT2C−/− mice displayed a reduced seizure threshold and a more rapid progression through the tonic-clonic phase of the seizure. The unexpected susceptibility of 5-HT2C−/− mice to epileptic seizures raises the possibility that this receptor participates in the modulation of neuronal network excitability. The ability to affect the seizure characteristics of normal mice with 5-HT2C ligands suggests that the phenotype of the 5-HT2C−/− mice is not a result of chronic compensation by other receptors for the loss of 5-HT2C receptors.
5-HT2C−/− mice were also noted to be significantly heavier than matched wild-type controls. Close analysis of their feeding behavior indicated that the weight increase was due entirely to increased intake, and not to a metabolic derangement that increases the efficiency of caloric storage. The serotonergic agonist 1-(3-chlorophenyl) piperazine (mCPP) drastically reduced food intake by wild-type mice, but had no effect on food intake by 5-HT2C−/− mice, demonstrating that appetite suppressants may operate via 5-HT2C receptors.
-eg